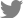Introduction
Toxicity testing is necessary to evaluate the potential risks to humans and animals posed by industrial and consumer products, as well as environmental factors. To date, a wide variety of animals have been sacrificed instead of humans to investigate biological mechanisms under toxic conditions. However, as limitations regarding ethical issues and species differences in animal experiments have been pointed out, alternatives have strongly been recommended. Although toxicity testing has commonly been conducted based on 2-dimensional (2D) culture in vitro, this method is unable to adequately represent the mechanisms of cell-cell and cell-matrix interactions and actual human organ functions. Given these limitations, organoid technology can be considered as a new alternative to animal experiments. Therefore, 3-dimensional (3D) culture systems have been proposed as a method to recapitulate the physiological conditions and architecture of human organs better than 2D culture systems [1].
Since the human lung is composed of multiple cell types, including epithelial cells, endothelial cells, stromal cells, and immune cells, various types of cells are needed to recapitulate the pulmonary environment. Alveolar type II epithelial cells (AEC2s) are widely known to be essential for alveolar renewal, repair, homeostasis, and maintenance [2-4]. In fact, AEC2s are capable of differentiating into alveolar type I epithelial cells (AEC1s), which cover 95% of the alveolar surface area and are involved in gas exchange during respiration [5,6]. We have previously shown that our protocol for obtaining alveolar organoids (AOs) derived from human pluripotent stem cells can generate multicellular AOs containing alveolar progenitor cells (EPCAM+CPM+NKX2.1+ cells), AEC1s (AQP5+ cells), AEC2s (SFTPC+ cells) and mesenchymal cells (VIM+ cells) [7]. In this study, we aimed to develop an assay for the assessment of pulmonary toxicity based on our 3D organoid culture method in vitro.
Nitric oxide (NO) plays an important role in physiological processes and mediating the pathogenesis of infection and reactive free radicals produced by inflammatory cells [8-10]. A previous study demonstrated that human lung epithelial cells produce NO in response to cytokine stimulation [11]. Therefore, the NO assay is an easy, effective, and indirect tool that could be used to evaluate the level of damage caused by inflammatory injuries.
In this study, we exposed AOs to 2 harmful chemicals, acrolein and sodium chromate, observed physical changes and NO responses at different times, and proposed a method for measuring cytotoxicity based on cell viability.
Materials and Methods
Ethics statement: Human induced pluripotent stem cells (hiPSCs) (C3) purchased by Korea Research Institute of Bioscience and Biotechnology (KRIBB, South Korea) were kindly provided. Therefore, approval by the institutional review board is not required.
1. Cell culture
Human induced pluripotent stem cells (hiPSCs) (C3) were kindly provided by Korea Research Institute of Bioscience and Biotechnology (KRIBB, South Korea) and cultured under xeno-, serum-, and feeder-free conditions. Cells were incubated in TeSR E8 basal medium (STEMCELL Tech, Vancouver, BC, Canada) on vitronectin-coated dishes. The hiPSC colonies were passaged by mechanical dissociation and transferred to new vitronectin-coated dishes with E8 medium containing Y-27632, a ROCK inhibitor. After overnight culturing, the medium was changed without a ROCK inhibitor to maintain the undifferentiated condition of the hiPSCs.
2. Generation of AOs from hiPSCs
The protocol for the generation of AOs was performed as reported in a previous study, with minor modifications [12]. First, hiPSCs were seeded with 10 to 20 small pieces from colonies with TeSR E8 basal medium on vitronectin-coated plates. When the colony size expanded to approximately 500 μm in diameter, the culture medium was replaced with differentiation medium to mediate definitive endoderm formation: RPMI-1640 containing 100 ng/mL activin A, 1 μM CHIR990212, 2X B27 in RPMI 1640 and 1% p/s as basal medium (day 0); 0.25 mM sodium butyrate in basal medium (day 1); and 0.125 mM sodium butyrate in basal medium (days 2-6). The basal medium from day 6 to day 21 was DMEM/F12 medium containing 0.05 mg/mL L-ascorbic acid, 0.4 mM monothioglycerol, 2X B27, and 1% p/s. On days 6 to 10, basal medium with 100 ng/mL noggin and 10 μM SB431542 was added to the cells to induce anterior foregut endoderm. On days 10 to 14, basal medium with 20 ng/mL BMP4, 0.5 μM ATRA, and 3.5 μM CHIR99021 was added to the cells to ventralize the anterior foregut endoderm. Cells were detached on day 14 of iAT2 cell differentiation with 0.4 U/mL collagenase B for 20 minutes and dissociated with enzyme-free cell dissociation buffer for 20 minutes. Next, cells were seeded into Corning 96-well round-bottom ultra-low-attachment microplates (Corning, Corning, NY, USA) at a seeding density of 7×103 cells/well in 100 μL. The round-bottom plate was centrifuged for 4 minutes at 1,500 rpm, and immediately, 50 μL of Matrigel solution diluted 1:7 in culture medium was carefully added to each well. After overnight culture at 37°C, the aggregated cells were transferred into Corning 96-well flat bottom ultra-low attachment microplates with new differentiation medium and cultured for day 24. On days 14 to 21, basal medium with 10 ng/mL FGF10, 3 μM CHIR99021, 10 ng/mL KGF, and 20 μM DAPT was added to the cells to induce alveolar progenitor cells. To differentiate alveolar and distal airway epithelial cells, DMEM Ham’s F12 medium containing 0.25% bovine serum albumin, 0.1% ITS, 1X B27, 50 nM dexamethasone, 0.1 mM IBMX, 100 ng/mL KGF, and 1% p/s was added to the cells on days 21 to 25 (Fig. 1). The culture medium was replaced every 2 days.
3. Chemical exposure of AOs
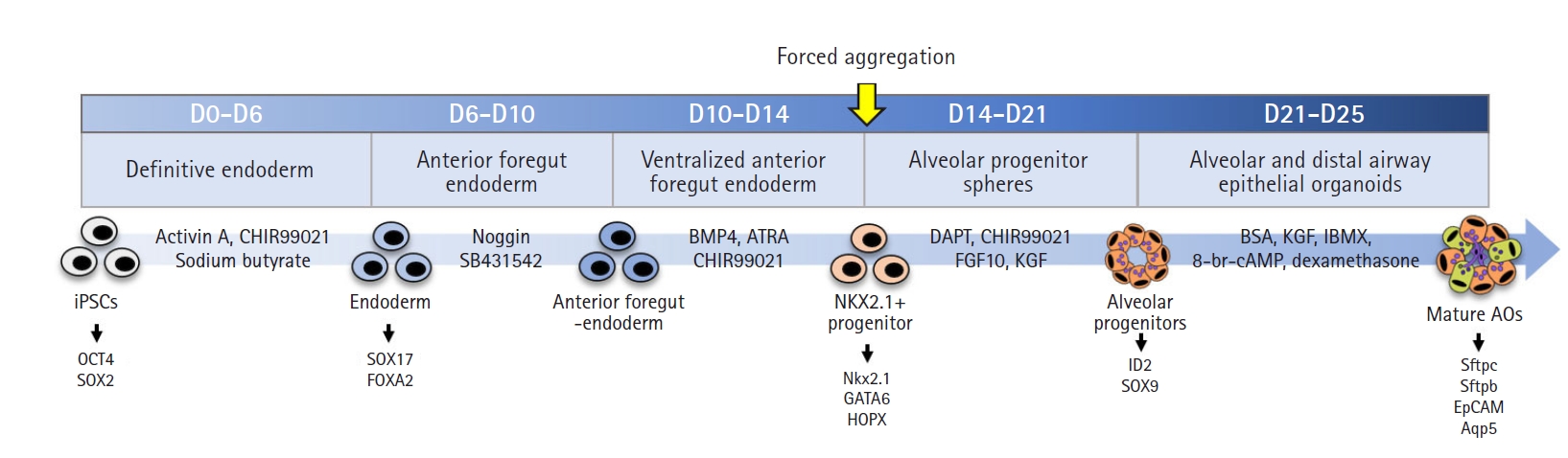
The AOs were exposed to acrolein (Cat. 110221; Sigma, St. Louis, MO, USA) and sodium chromate (Cat. 307831; Sigma) on day 20 (for 120 hours of exposure), day 22 (for 72 hours of exposure) and day 24 (for 24 hours of exposure) until day 25. Acrolein was diluted to concentrations of 0 to 1,280 μg/mL in phosphate-buffered saline (PBS). Sodium chromate was diluted to concentrations of 0 to 100 μg/mL) in PBS. Organoid size was estimated under a microscope.
4. MTT assay
AOs were exposed to acrolein and sodium chromate for 24, 72, and 120 hours. On the final exposure day, 0.1 mg/mL MTT reagent (thiazolyl blue tetrazolium bromide) (Cat. m5655; Sigma) was added to the plate wells for 2 hours in a 37°C incubator. The reagent was clearly removed, and then formazan crystals were dissolved in dimethyl sulfoxide. The absorbance was measured at 540 nm using a microplate reader, and cell viability was calculated as follows: cell viability (%)=(Atreatment-Ablank)/(Acontrol-Ablank)×100%, where A indicates the absorbance in terms of optical density.
5. Nitric oxide assay
The amount of extracellular nitrite release in the cell supernatant was measured by a modified Griess reagent assay (Cat. G4410; Sigma). Fifty microliters of Griess reagent and an equal volume of cell supernatant were immediately reacted for 7 minutes at room temperature, and absorbance was measured at 540 nm on a microplate reader. The level of NO was calculated using an equation derived from linear regression analysis using a standard curve of sodium nitrite solution.
6. Statistical analysis
The results are represented as the mean±standard error of the mean. One-way analysis of variance followed by the Bonferroni multiple comparison test was used for comparisons between multiple groups. The concentration responsible for half-maximal cell viability (IC50) was calculated using GraphPad Prism 9 software (GraphPad Software, San Diego, CA, USA). A p-value <0.05 was considered to indicate statistical significance.
Results
1. 3D organoid-based toxicity assays at different exposure times
We generated AOs to develop a tool to assess chemical toxicity in vitro. AOs were observed with empty regions that resembled alveolar sacs, and they were composed of well-arranged cells on their surface (Fig. 2, control). However, the damaged cells were detached from the AOs’ surface after exposure to acrolein or sodium chromate, generating a surface line of AO structure that looked ambiguous compared with the control AOs (Fig. 2). At 24 hours after exposure, cell viability decreased in a dose-dependent manner from acrolein concentrations above 5 μg/mL and sodium chromate concentrations above 3.13 μg/mL (Fig. 3A and 3D) in MTT assay. Since the alveolar structure was disrupted with a loss of cells through cell death, we anticipated that chemical exposure would lead to a decrease in organoid size. However, organoid size did not change significantly after 24 hours (Fig. 3B and 3E, 24-hour group).
Subsequently, we exposed the AOs to chemicals for longer than 24 hours (72 hours and 120 hours) to validate whether toxic stimulation-induced cell loss could affect organoid size. After 72 hours of exposure, cell viability was similar to that observed after 24 hours of exposure (Fig. 3A, 72-hour group). However, a high dose of acrolein (above 40 μg/mL) led to a reduction in organoid size (Fig. 3B, 72-hour group). AOs treated with sodium chromate seemed to decrease in size, but not significantly (Fig. 3E, 72-hour group). After 120 hours of exposure, cell viability decreased from a low-dose treatment.
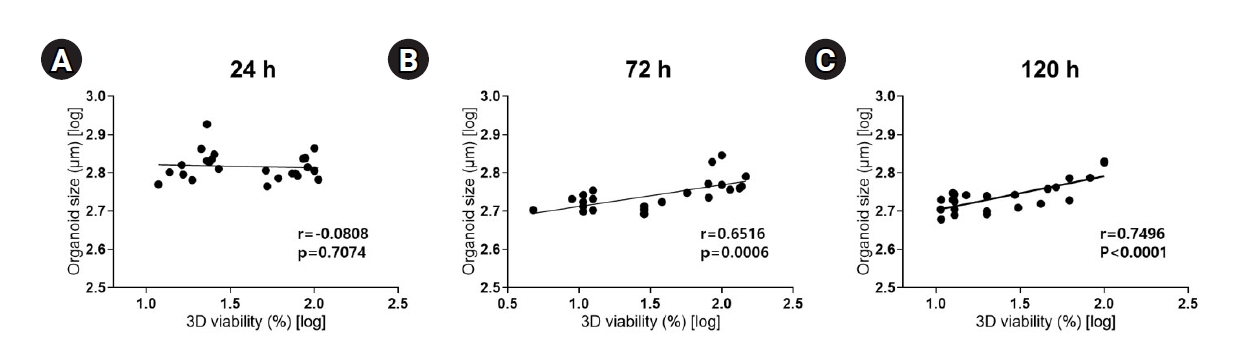
2. Correlation between organoid size and viability
We sought to verify a relationship between organoid size and viability depending on exposure time. After 24 hours of exposure to acrolein and sodium chromate, the correlation coefficient between size and viability was calculated to be r=-0.0808, indicating no meaningful correlation (Fig. 4A). After 72 hours of exposure, the correlation coefficient was r=0.6516, indicating a strong positive correlation (Fig. 4B). After 120 hours of exposure, the correlation coefficient was r=0.7496, also indicating a high positive correlation (Fig. 4C). These data showed that correlation between organoid size and viability increased with a longer exposure time. Taken together, a comparative analysis of organoid size after a toxic injury is suitable for toxicity testing after at least 24 hours of exposure.
Discussion
In this study, we performed toxicity assays using 2 chemicals: acrolein and sodium chromate. Typically, cell viability assays provide a readout of cell health through measurements of ATP content or cell proliferation in 2D culture [13]. In this study, we conducted the MTT assay, which has been widely used to measure cell viability and cytotoxicity [14]. Another cell proliferation assay based on colorimetric analysis, the WST-1 (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt) assay, forms water-soluble formazan dye, unlike the MTT assay, which forms insoluble purple formazan crystals [15]. We observed that the WST-1 assay generated inaccurate data with large variations (data not shown). Therefore, we suggest that the MTT assay is suitable for measuring cell viability of organoids on a 96-well plate template rather than a water-soluble formazan dye-based colorimetric assay. In addition, several studies have suggested that organoids damaged by exposure to some stimuli revealed reductions in size. For example, infection of brain organoids with Zika virus resulted in a reduction in organoid size [16]. Moreover, Moll et al. [17] reported the toxicity of Cre-recombinase, which led to a reduction of diameter in enterocyte organoids. They stated that the size of organoids was affected by reduced cell proliferation, induction of apoptosis, and DNA damage. Accordingly, we suggest that organoid size could be used as an indirect indicator of toxicity, as necrotic or apoptotic epithelial cells are shed from organoids, leading to a decrease in organoid size. Although cell viability decreased in response to short-term exposure (less than 24 hours), the organoid size did not change, and no correlation was found between size and viability. However, long-term exposure to chemicals (over 72 hours) induced high cytotoxicity, with a decrease in cell viability, and a high positive correlation was observed between size and viability under those conditions. Taken together, it is possible to establish a toxicity assessment platform based on our AOs by performing the MTT assay in short- and long-term exposure models. Furthermore, organoid size could be a useful indicator of toxicity in long-term culture, rather than in short-term culture (less than 24 hours).
NO is commonly associated with various pulmonary diseases, including chronic obstructive pulmonary disease, idiopathic pulmonary fibrosis, and acute respiratory distress syndrome/acute lung injury [18-20]. Under inflammatory conditions, NO is synthesized in many cell types, accompanied by cell death and apoptosis [21,22]. Therefore, NO synthesis has been considered as a toxic response after an inflammatory injury [23]. Likewise, alveolar epithelial cells also produce NO in response to signaling from the innate immune system in a toxic environment [24]. In a previous study, we demonstrated that cigarette smoke extract injury induced NO production in a mouse type II epithelial cell line, MLE-12, and a human type II epithelial cell line, A549 [18,25]. In this study, acrolein and sodium chromate induced NO production in AOs at high doses. Furthermore, the NO level responded more sensitively to chemical exposure in a time-dependent manner. These data indicate that NO might be a tool that could be used to assess toxicity in 3D culture. Since NO is typically produced by inflammatory cells including macrophages and neutrophils [26,27], the presence of inflammatory cells with AOs might increase the NO response to injury and better recapitulate the human physiological environment.
In summary, we generated AOs to develop a toxicity screening tool. Our 3D organoid technology is a valuable model system to measure cytotoxicity based on a cell viability assay in vitro. Although it is difficult to compare the size variation of organoids in acute toxicity testing under 24 hours of exposure, cell viability was an effective indicator of cytotoxicity. After long-term exposure, however, both cell viability and organoid size could be tools to evaluate the toxicity of various hazardous materials. This method might be especially useful for assessing toxicity in the respiratory system.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print