Introduction
Intestinal epithelial cells (IECs), which comprise functional cells such as goblet cells and Paneth cells, play a crucial role in maintaining intestinal homeostasis [1,2]. In addition to their essential functions such as absorption of water and nutrients, IECs serve as physical and chemical barriers, protecting the internal environment from symbiotic microbial insults [1,3,4]. To sustain these barrier functions, IECs engage in continuous replenishment, originating from intestinal stem cells that undergo self-renewal every 4 to 5 days [3]. During the process of self-renewal in IECs, Lgr5+ stem cells differentiate into transit-amplifying (TA) cells [5]. Subsequently, these TA progenitor cells sequentially adopt the fates of secretory cells, such as goblet, Paneth, and enteroendocrine or absorptive enterocyte cells [5]. Therefore, maintaining the proper balance of IECs is essential for preserving normal intestinal function. The intestine is a structurally complex organ that is suitable for reproduction through the organoid technique [6,7]. In addition, colon organoids can replicate the typical histological features of stem cell-based organs [8]. With their capacity for functional cell differentiation, colon organoids are extensively used in various research fields, including studies of the microbiome, disease mechanisms, and drug discovery [8-11].
Leucine-rich repeat-containing G-protein-coupled receptor 5 (Lgr5) is a G protein-coupled receptor (GPCR) and a stem cell marker recognized in various organs, including the small intestine [12]. Furthermore, Lgr5 recognizes R-spondins on the cell membrane [13] and is associated with the initiation of the Wnt/β-catenin signaling pathway and regulation of tissue homeostasis and stem cell biology [14,15]. Conversely, a decrease in Lgr5 expression under endoplasmic reticulum (ER) stress is observed in human colorectal cancer [16]. In particular, protein kinase RNA-like ER kinase (PERK) is more closely related to Lgr5 depletion than other ER-sensing proteins, such as inositol-requiring enzyme 1 (IRE1) and activating transcription factor 6 (ATF6) [16]. In a previous study, ER DnaJ domain-containing protein 5 (ERdj5) deficiency increased PERK-related ER stress under inflammatory conditions, leading to cell apoptosis [17]. However, the correlation between ERdj5 and Lgr5 expression in the intestinal epithelium remains unclear.
The ER DnaJ domain-containing protein 5 (ERdj5), a disulfide reductase in the ER, moderates the processing of misfolded proteins [18]. The accumulation of unfolded or misfolded proteins inside the ER due to various factors induces stress [19,20]. Moreover, ER stress is caused by excessive nutrient accumulation, irradiation, and infection and is known to be especially related to inflammation [21-23]. In a previous study, it was established that while ERdj5 does not pose any issues concerning the growth of IECs, the protein leads to excessive ER stress and subsequent apoptosis in certain specialized cells, particularly goblet cells, when exposed to inflammatory conditions [17]. Additionally, irradiation induces ER stress by producing reactive oxygen species (ROS) [24]. Furthermore, ER signaling proteins such as ATF6, PERK, and IRE1 are activated by irradiation-induced ER stress. Autophagy, the DNA damage response, and the ROS stress response are affected by ER stress [25].
In this study, we cultured colon organoids (colonoids) using a crypt obtained from the colon epithelium and hypothesized that the ER-related protein ERdj5 affects proliferation markers in IECs.
Materials and Methods
Ethics statement:
All experiments were performed following the guidelines and protocols approved by the Institutional Animal Care and Use Committee of Kangwon National University (IACUC, KW-210610-5).
1. Mice
Female C57BL/6 mice and ERdj5 deficient mice were purchased as previously described [17]. All the mice were bred at the Kangwon National University Animal Laboratory Center.
2. Organoid culture
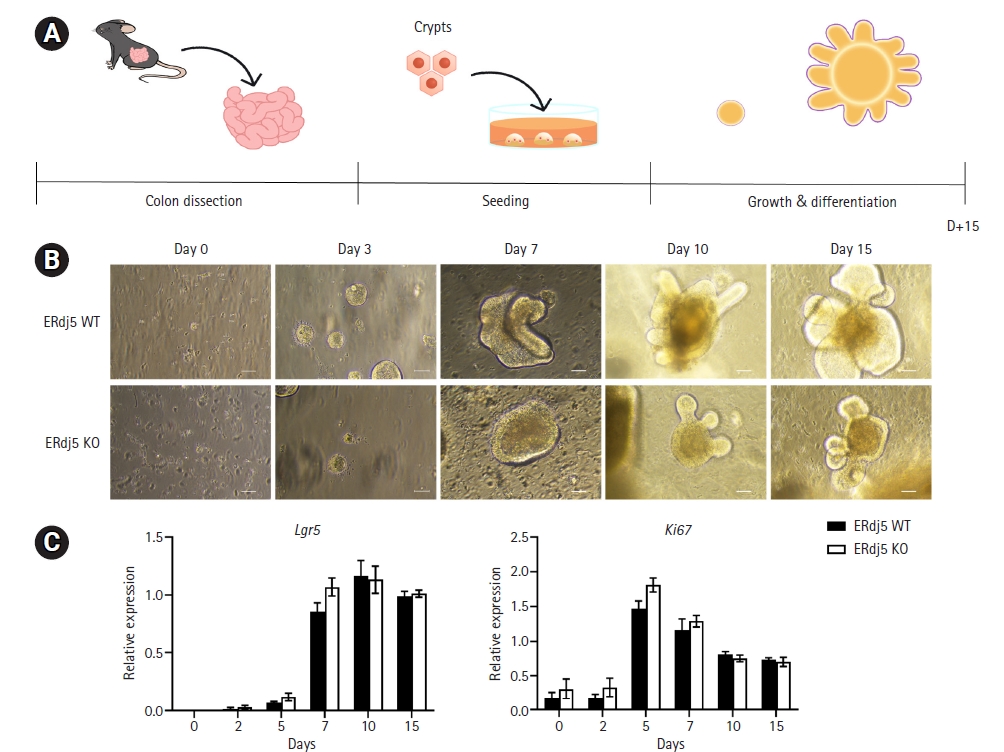
Murine colonic organoids were cultured as previously described [26]. Briefly, mouse colonic tissues were washed to eliminate the gut microbiota, and dissociated colonic crypts were used as a recovery solution. Finally, intestinal crypts were plated in 50% Matrigel and filled with colon organoid growth medium (Dulbecco's Modified Eagle Medium F/12; Gibco, Carlsbad, CA, USA), 50% L-WRN growth medium, B27 (Gibco), N2 (Gibco), 1.25 mM N-acetylcysteine (Sigma Aldrich, St. Louis, MO, USA), 10 mM nicotinamide (Sigma Aldrich), 10 μM P38i (Sigma Aldrich), 16.7 pM mEGF (Invitrogen, Vienna, Austria), 10 μM Y-27632 (Abmole, Houston, TX, USA), and 0.5 μM A83-01 (Tocris, Bristol, UK). The colon organoid growth medium was replaced every 2 days, and the colonoids were passaged every 7 days. Colon organoids were observed using Nikon Eclipse Ts2 (Nikon, Minato, Tokyo, Japan).
3. Organoid stress induction
Colonoids were incubated for 48 hours at 37°C on day 7 with 1 µg/mL of Pam3CSK4 (InvivoGen, San Diego, CA, USA), an agonist of Toll-like receptor (TLR)2/TLR1, to mimic inflammatory conditions.
Radiation exposure experiments inducing cell stress in colonoids were performed as previously described. In brief, we treated the colonoids after 3 days of culture with a 6 Gy dose of γ-radiation using Gammacell 40 Exactor (Best Theratronics, Ottawa, ON, Canada). After radiation treatment, the colonoids were incubated at 37℃ for 4 days.
4. Quantitative real-time polymerase chain reaction
Total RNA was extracted from the colon organoids using the TRIzol reagent (Ambion, Austin, TX, USA). RNA quality was confirmed using an ultraviolet-visible spectrophotometer (MicroDigital, Seongnam, Korea). The RNA was reverse-transcribed using the Goscript Reverse Transcription System (Promega, Madison, WI, USA). Additionally, quantitative real-time polymerase chain reaction (qRT-PCR) was performed using GoTaq® qPCR and RT-qPCR Systems (Promega) by CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The primers used for qPCR were in Table 1.
5. Immunofluorescence staining
Colon organoids were fixed in 4% paraformaldehyde and permeabilized with 2% Triton-X100 (Sigma Aldrich). Nonspecific antibody binding to the organoid surface was blocked with 3% bovine serum albumin (MP Biomedicals, Solon, OH, USA) in phosphate-buffered saline. To assess colon cell death, the ApopTag Fluorescein In Situ Apoptosis Detection Kit (Merck, Kenilworth, NJ, USA) was used for the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay. The colonoids were then mounted with a Fluoroshield with 4′,6-diamidino-2-phenylindole (Sigma) to visualize the nuclei. Organoid images were visualized using a confocal laser scanning microscope (LSM880; Carl Zeiss, Göttingen, Germany) and analyzed using the ZEN program (Carl Zeiss).
6. Statistical analysis
GraphPad Prism 10 (San Diego, CA, USA) was used for data analysis. Differences between the 2 groups were analyzed using the t-test. One-way analysis of variance and post-hoc tests were also performed. The Tukey multiple-comparison test was utilized for comparisons of more than 2 groups. A threshold of p<0.05 was deemed statistically significant.
Results
1. Phenotype changes in the in vitro developmental process of colon organoids
We cultured an equal number of crypts (5×102/well) from the ERdj5 wild-type (WT)/ knockout (KO) mice colon and changed the medium every 2 days while observing the growth of the organoids for 15 days (Fig. 1A). We observed the morphology of the organoids at the indicated time points and obtained RNA to observe changes in gene expression (Fig. 1B). As observed in the colonoid images, no apparent differences in the early stages of growth were noted; however, from day 7, the ERdj5 KO organoids were smaller than the WT organoids. Moreover, Lgr5 levels on the indicated dates rapidly increased until day 7 in both the ERdj5 WT and KO groups, followed by a significant decrease thereafter (Fig. 1C). However, no significant difference in Lgr5 and Ki67 expression was observed between WT and ERdj5 KO organoids (Fig. 1C). We investigated differentiation because we did not observe significant differences in growth factors between ERdj5 WT and KO colon organoids over 15 days.
2. Genetic changes associated with differentiation
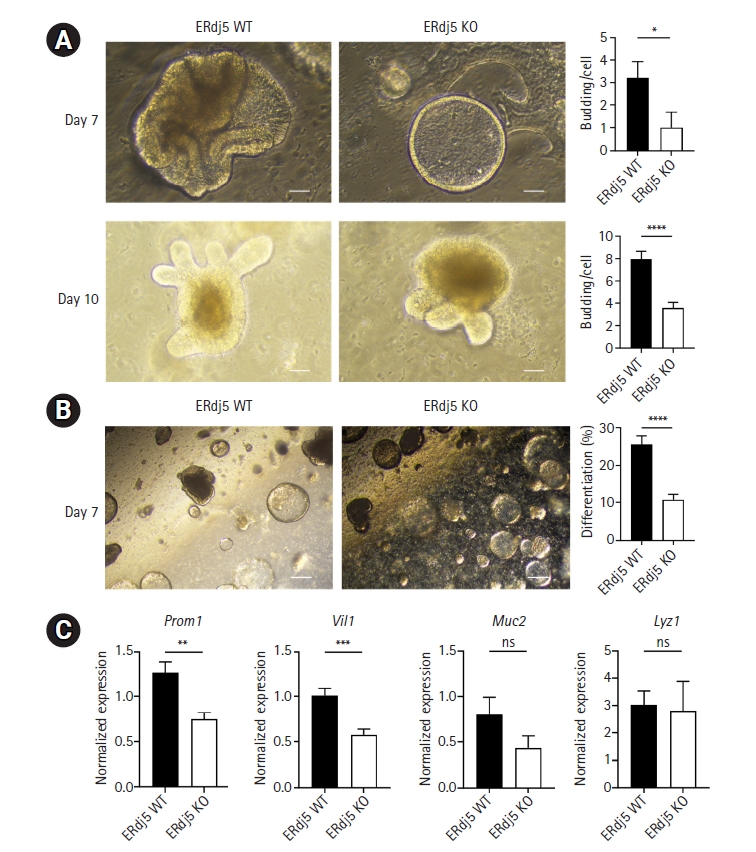
Previous studies have confirmed the appearance of differentiation-related characteristics that confer colonic identity on days 7 and 10 [27]. To evaluate the organoids, we analyzed budding on days 7 and 10. We observed that ERdj5 KO organoids formed fewer complex crypt-like structures than WT organoids (Fig. 2A). Furthermore, when evaluating the degree of differentiation on day 7 using microscopy, we observed that the level of differentiation per well was less pronounced in ERdj5 KO mice than in WT mice (Fig. 2B). Next, we confirmed the presence of RNA in functional cells that constitute the colon. The mucin 2 (Muc2) marker in goblet cells and lysozyme 1 (Lyz1) marker in Paneth cells demonstrated no significant differences between the groups. However, the enterocyte marker Villin 1 (Vil1) and the TA cell marker Prominin 1 (Prom1) exhibited lower levels in ERDj5 KO colonoids on day 7 than in WT colonoids (Fig. 2C). Collectively, the absence of ERdj5 resulted in reduced organoid differentiation.
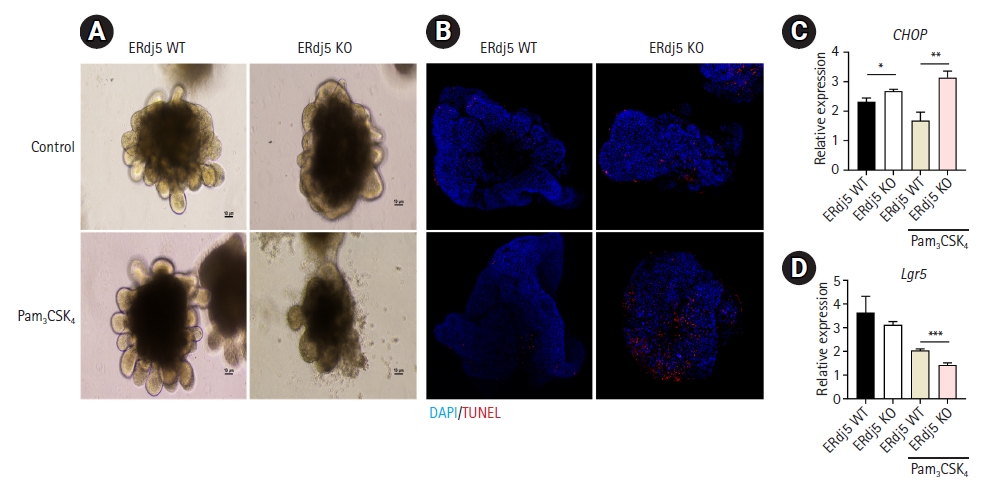
3. Comparison of changes in response to Pam3CSK4-induced inflammatory stimulation between ERdj5 WT and ERdj5 KO organoids
ER stress modulates the levels of Lgr5, a prominent stem cell marker and a key factor in organoid growth [28]. As reported in our previous study, we induced inflammation using Pam3CSK4 (a TLR2 ligand) and observed external structural collapse, increased C-X-C motif chemokine ligand 1 secretion, and goblet cell depletion [17]. We investigated the impact of ERdj5 on functional cells such as Paneth cells and goblet cells in IECs; however, the influence of ERdj5 on the differentiation of stem cells into functional cell types remains unknown. Therefore, we aimed to investigate the effect of ERdj5 deficiency on growth factors in differentiated organoids by inducing TLR2-mediated inflammation. After 7 days of growth, colonoids were incubated for 48 hours at 37°C with 1 µg/mL of Pam3CSK4, which led to structural rupture in ERdj5-deficient organoids, unlike WT organoids (Fig. 3A). Similarly, as displayed in Fig. 3A, ERdj5 deficiency increased the number of TUNEL-positive cells (Fig. 3B). We examined various inflammation-related factors at the RNA level; however, no significant differences were identified except in interleukin-1 beta (Fig. S1). However, an increase in the PERK-related protein C/EBP homologous protein (CHOP) provided insight into the occurrence of cell apoptosis (Fig. 3C). Additionally, induction of ER stress was associated with a significant decrease in Lgr5 expression (Fig. 3D). Pam3CSK4 treatment induced ER stress, resulting in the exacerbation of apoptosis and stemness in organoids with ERdj5 deficiency.
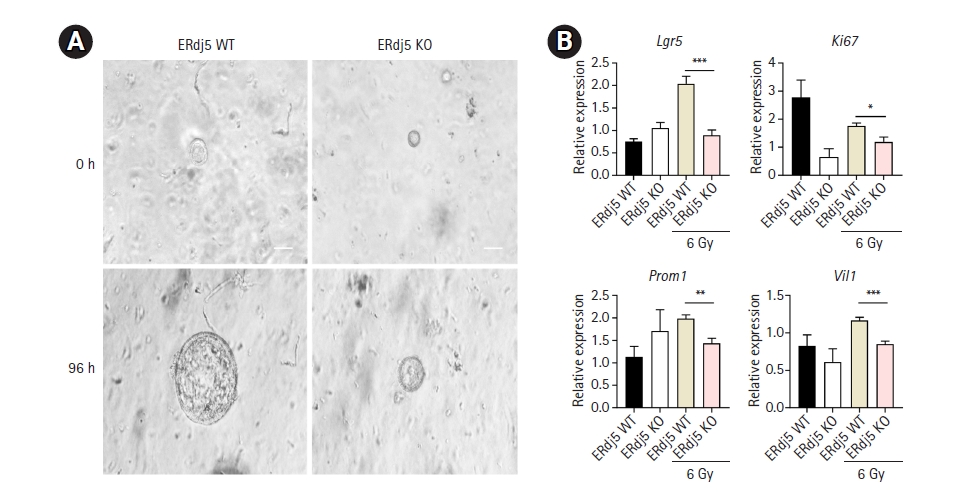
4. Comparison between WT and ERdj5 KO organoids concerning ROS-associated inflammation
Irradiation induces ER stress by directly or indirectly generating ROS [25]. We induced ER stress through another pathway to investigate how ERdj5 affects organoid growth. A previous study found that radiation with an intensity greater than 6 Gy resulted in the loss of budding and morphology maintenance in organoids [29]. Following this reference, we cultured organoids for 3 days and then irradiated them with 6 Gy to investigate the changes occurring. In ERdj5-deficient organoids, no expansion was observed, resulting in smaller organoids than the WT organoids (Fig. 4A). In WT organoids, Lgr5 exhibited a significant increase 4 days after irradiation, whereas, in ERdj5 KO organoids, no significant increase was observed (Fig. 4B). After irradiation, the markers Prom1, Vil1, and Ki67 decreased in ERdj5 KO organoids compared to WT organoids, indicating a decline before their differentiation into functional cells from stem cells. However, no significant differences were observed in the ER stress initiator binding immunoglobulin protein (BiP) after irradiation. Collectively, inflammation through irradiation induced mild ER stress in both groups, whereas ERdj5 deficiency eventually led to the downregulation of Lgr5. The colonoids were unable to defend against various inflammatory stimuli owing to a deficiency of the ERdj5-related ER stress resolution pathway.
Discussion
The growth of organoids follows a sequence of processes, including the collection of initial cells and stem cells, proliferation, migration, and finally differentiation [8]. Although we observed no morphological differences up to day 5 in previous studies, we did not investigate the changes that occurred within the organoids during differentiation.
In this study, we identified genetic changes associated with the proliferation and differentiation of IECs in mouse colon organoids. No significant differences were identified in proliferation-related markers, such as Lgr5 and Ki67, under steady-state conditions. However, ERdj5-deficient organoids reduced the expression of the epithelial marker Vil1 and the TA cell marker Prom1. Moreover, no noticeable differences were observed in the expression of the aforementioned markers, Muc2 for goblet cells and Lyz1 for Paneth cells. However, additional research is necessary to explore potential changes in other functional cell genes.
Additionally, ER stress induces stemness in IECs [30]. We utilized the TLR2 ligand Pam3CSK4 and irradiation as pathways for inducing ER stress. In our previous study, we identified that TLR2 treatment led to preferential apoptosis of goblet cells [17]. In this study, we observed increased apoptosis and CHOP gene expression in ERdj5 KO colonoids. Furthermore, we observed a significant decrease in Lgr5 following Pam3CSK4 treatment. Increases in the expression of various inflammatory genes were observed; however, no significant differences were present between the WT and ERdj5 KO colonoids. Radiation exposure induces ER stress by triggering intracellular ROS [25]. In a previous study, exposure of organoids to a radiation dose of 6 Gy did not exhibit significant effects on organoid growth [29]. We did not observe any differences in the expression of the ER stress initiator gene BiP after radiation exposure. However, Prom1 and Lgr5 were expressed at significantly lower levels in ERdj5 KO mice than in WT mice after irradiation. The absence of ERdj5 appeared to lead to growth issues in organoids following irradiation.
Colon organoids must maintain their proper growth and differentiation [8]. If homeostasis is not sustained, cells may undergo apoptosis [31]. During the early stages of colon organoid development, the presence of ERdj5 is not considered crucial. However, as differentiation is initiated, ER stress appears to render the maintenance of homeostasis more vulnerable. Although additional research is necessary, our findings highlight the importance of ER stress regulation in colon organoid growth. This indicated that the absence of ERdj5 under stress could make colon organoids more susceptible to differentiation and growth challenges.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement
Supplement Print
Print