Introduction
Organelles use distinct sub-compartments to spatially organize their biological reactions. Unlike membrane-bound organelles that have a lipid membrane, non-membrane-bound organelles form distinct compartments through a process called liquid-liquid phase separation (LLPS) due to the lack of lipid barriers [1]. In this process, a homogeneous solution is separated into 2 distinct liquid phases. This results in highly condensed RNAs and RNA-binding proteins (RBPs) that function as single organelles [2]. These aggregates are a hallmark of several neurodegenerative diseases, including ParkinsonŌĆÖs disease, amyotrophic lateral sclerosis (ALS), and frontotemporal dementia (FTD) [3].
The dynamics of the condensates of membrane-less organelles provide multiple strategic advantages over membrane-bound organelles because they can condense macromolecules and rapidly exchange constituents with the neighboring cytoplasm or nucleoplasm [4]. An example is furnished by the numerous RNA-protein condensates, termed ribonucleoprotein (RNP) granules, which are dynamic and assembled by LLPS [5]. In eukaryotic cells, RNP granules exist in the nucleus (Cajal bodies, nucleoli, and paraspeckles) and cytosol (processing bodies and stress granules [SGs]) [6-8]. SGs are well-known RNP granules, and many studies on them have recently been reported.
SGs are membrane-less cytoplasmic aggregates of mRNAs and associated proteins in eukaryotes with a size of 0.1 to 2 ┬Ąm and are formed under various environmental stresses such as oxidative stress, viral infection, hypoxia, heat shock, or ultraviolet (UV) irradiation [9,10]. SGs include stalled translation initiation complexes such as ribosomal subunits, mRNA molecules, and diverse RBPs [11,12]. SG assembly leads to translational arrest and formation of preinitiation complexes (PICs), which consist of mRNAs, ribosomal subunits, and related translation initiation factors and are distributed heterogeneously within the granules [13,14].
Although animal models have been widely used in research, they have an inherent limitation of physiological differences between humans and animals. Organoids have recently emerged as useful in vitro models that recapitulate aspects of organs, such as being composed of various tissue-specific cells, self-organizing spatially, and having tissue-specific functions [15-18]. Therefore, they are useful for disease modeling, development research, drug screening, regenerative medicine, and toxicity studies. This review examines the mechanism of SG formation and discusses their roles in various human diseases, particularly lung disease. Finally, we discuss the potential of SG-associated proteins to be widely utilized as markers for lung toxicity assessment using lung organoids (LOs).
Mechanism of SG formation
According to several proposed mechanisms of SG assembly, the distinctive interaction of mRNAs with RBPs inhibits the translation process of protein synthesis for specific mRNAs by altering the activity of the eukaryotic initiation factors eIF2╬▒ or the eIF4F complex. Type I SGs are formed under conditions of endoplasmic reticulum stress, oxidative stress, or viral infection via phosphorylation of components of the eIF2╬▒ pathway. Type II SGs are phospho-eIF2╬▒-independent and are primarily formed through the inactivation of eIF4A. Type III SGs are triggered by UV light and glucose starvation and lack eIF3 [19-23]. Among them, the most well-studied pathway is one in which triggering eIF2╬▒ phosphorylation causes the subsequent aggregation of stalled translation initiation factors (Fig. 1). First, translation initiation is inhibited, which results in the accumulation of stalled mRNAs. In most cases, translational arrest occurs through phosphorylation of eIF2╬▒ by kinases such as heme-regulated inhibitor (HRI), protein kinase R (PKR), PKR-like endoplasmic reticulum kinase (PERK), and general control nonderepressible 2 (GCN2). Second, the stalled mRNAs and associated proteins are condensed and form distinct cytoplasmic foci. The phosphorylation of eIF2╬▒ disrupts translation initiation and, consequently, 48S PICs cannot recruit the 60S ribosomal subunit, resulting in stalled 48S PICs being recruited to the SGs [24,25].
The assembly and disassembly of SGs are regulated by numerous RBPs. SG-related proteins have been studied over the past 10 years, and more than 150 proteins have been identified, of which 3 fundamental components are poly(A)-binding protein (PABP), T-cell intracellular antigen-1 (TIA-1), and Ras GAPSH3-binding proteins (G3BPs) [26,27]. PABP, TIA-1, and G3BPs are RBPs that regulate mRNA translation and stability. PABP is a protein generally diffused in the cytoplasm that forms discrete foci in response to stress. TIA-1 co-localizes with TIA-1-related protein (TIAR) at SGs, resides in the nucleus, and rapidly accumulates in the cytoplasm under cellular stress. Although PABP and TIA-1 are core components of SGs, they are not essential for SG assembly because when these genes are knocked out, SG formation is either reduced or delayed, but not completely halted [28]. G3BPs (G3BP1 and G3BP2), which are the most well-studied RBPs, consist of 2 globular domains: the nuclear transport factor 2-like (NTF2) domain on the N-terminus and the RNA recognition motif (RRM) on the C-terminus, with intrinsically disordered regions [29,30]. G3BPs are considered to be molecular switches that regulate RNA-dependent LLPS. A study demonstrated that G3BPs are the central nodes of the network between core proteins and RNA, resulting in SG assembly through LLPS [28]. Moreover, several studies have demonstrated that G3BPs are essential for SG formation, which is initiated by the phosphorylation of eIF2╬▒ through PKR activity, based on the results that G3BP1 overexpression in somatic cells induced spontaneous SG condensation and, conversely, silencing G3BP1 inhibited SG condensation [31,32]. Both G3BP1 and G3BP2 are recruited to SGs and play essential roles in their formation in various human cells. According to Matsuki et al., while the knockdown of G3BP2 did not substantially reduce SG formation, the knockdown of G3BP1 strikingly affected SG formation in 293T and HeLa cells [33]. Therefore, G3BP1 rather than G3BP2 is commonly used as an SG marker [34].
SGs and human diseases
SGs have been associated with many diseases that are characterized as having toxic aggregates because LLPS tends to condense inherently into dangerous proteins [35,36]. Therefore, over the past several decades, many studies have been performed on the relationships of SGs with pathological aggregations and neurodegenerative disorders, such as Alzheimer disease, ALS, and FTD. Abnormal cytoplasmic aggregations of proteins, including TAR DNA binding protein 43 and fused in sarcoma (FUS), were detected in the neurons and glia of patients with ALS and FTD [37]. In addition, cytoplasmic aggregates of ubiquitin-associated protein 2-like, one of the SG components, have been detected in ALS [38]. SG formation by the accumulation of SG-related proteinsŌĆönamely, PABP, FUS, G3BP1, and TIA-1ŌĆöwas also observed under conditions of chronic stress with the neurodegenerative potential [26].
A recent study revealed that exposure to pesticides, such as N, N-diethyl-meta-toluamide, fipronil, or maneb, increases reactive oxygen species, and induces oxidative stress and SG formation via eIF2╬▒ phosphorylation, causing neurodegenerative disorders [39]. In addition, bisphenol A, a chemical produced during the production of polycarbonate plastics, can generate SGs by phosphorylation of eIF2╬▒ through the PERK pathway in human cells [40]. Collectively, SGs are closely associated with human diseases and play a vital role in defending host cells from harmful conditions.
Pulmonary diseases and oxidative stress
Lung disorders can be triggered by many factors, such as smoking tobacco, air pollution, silica dust, and viral infections. The relationship between pulmonary diseases and smoking was first recognized in the 1870s, and numerous studies have been conducted on this association. Cigarette smoke extract was found to trigger oxidative stress, leading to destruction of the alveolar wall and increased proinflammatory cytokines in the lungs of patients with chronic obstructive pulmonary disease (COPD) [41]. Similarly, nicotine, an addictive component of tobacco, was reported to be associated with COPD and lung cancer, as it induced the transformation of human bronchial epithelial cells [42].
Oxidative stress is involved in many biological processes and pulmonary diseases. Arsenite is one of the most common oxidative stress inducers, and a study has demonstrated that sodium arsenite (NaAsO2) causes cytotoxicity, apoptosis, genotoxicity, and oxidative stress in human lung cells [43]. Air pollutants, such as particulate matter and ozone, also cause oxidative stress. Moreover, these pollutants can cause inflammation, cellular damage, and impaired mitochondrial function in lung cells, leading to multiple respiratory diseases [44,45]. Air pollution is associated with several respiratory diseases, especially in patients with COPD, and the possibility of developing asthma-COPD overlap syndrome can increase at high concentrations of pollution [46,47].
These studies show that pulmonary diseases are associated with various risk factors and are driven by oxidative stress and that oxidative stress-inducing agents such as arsenite trigger SG formation. Arsenite-treated human epithelial cells form G3BP and TIA-1 granules, which are markers of SGs [48]. In addition, exposure to NaAsO2 induces the formation of G3BP1 granules, which are commonly used SG markers in human lung epithelial cells, including BEAS-2B and A549 cells [49]. Therefore, SGs may play a potential role as lung toxicity biomarkers because oxidative stress is one of the causes of SG formation.
Infectious lung diseases and SGs
Studies on infectious lung diseases have been extensively reported. A study demonstrated that patients with swine-origin influenza A virus infection developed pulmonary inflammation and lung injuries, fibrosis progression, and acute respiratory distress syndrome [50,51]. Additionally, several viruses lead to the formation of SGs, as exemplified by the formation of G3BP1-containing granules in dengue virus-infected A549 human lung epithelial cells [52]. The involvement of alphavirus virulence factor nsP3 in regulating the composition of biomolecular condensates, such as SGs, during alphavirus infection has also been reported [53].
Due to the coronavirus disease 2019 (COVID-19) pandemic, numerous studies have focused on infectious pulmonary diseases, especially respiratory illnesses [54,55]. Respiratory virus infections have been reported to induce considerable lung injuries and immune responses. Respiratory syncytial virus (RSV) is the most significant cause of acute respiratory infections in children and the elderly [56,57]. A previous study showed that RSV induces a stress response that continues throughout infection, indicating that RSV may specifically initiate and maintain stress responses [58]. Choi et al. recently reported that polyhexamethylene guanidine phosphate (PHMG-p), a humidifier disinfectant, induced SG formation in human lung epithelial cells upon RSV infections [49,59]. They also showed that PHMG-p treatment exacerbated cellular stress in RSV-infected cells, leading to DNA damage and eventually cell death [60]. Taken together, these results show the possibility that SG formation can be utilized as a marker for evaluating chemical toxicity, such as PHMG-p, as well as viral infections.
Lung organoid models
Numerous studies have reported on various organoids, including LOs [60-64]. Organoids can be derived from adult stem cells, which are tissue-specific pluripotent stem cells (PSCs), including induced PSCs and embryonic stem cells. Organoids using mouse tracheal basal cells expressing p63, Krt5, and Krt14 were generated by Rock et al. [64], and primary human airway basal cell-derived organoids consisting of ciliated cells, goblet cells, and basal cells were also generated [65]. Furthermore, PSC-derived LOs exhibiting gene expression patterns similar to those of human alveolar tissue have been generated [66]. In addition, the process of establishment of lung and airway organoids, such as Matrigel droplet culture or matrix-free suspension culture, has been well described [18,67,68]. We also generated human PSC-derived LOs in Matrigel droplets and confirmed that these organoids expressed SCGB3A2, which is a marker of secretory cells in the airway epithelium (Fig. 2).
Three-dimensional organoids can closely mimic functional characteristics and in vivo microarchitecture. Therefore, they are widely used to study development, disease modeling, and drug screening. To establish severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection models, human bronchial organoids were generated using bronchial epithelial cells [69] and a distal LO model containing alveolar type 2 and basal cells was established [70]. A study to verify the efficacy of therapeutic agents using human LOs derived from SARS-CoV-2-infected lung tissue was also reported [71].
In addition to SARS-CoV-2 infection models, human LO models for drug screening and toxicity evaluation of pharmaceuticals and chemicals have also been actively studied. Lung cancer organoid models for drug screening that can validate the responses of tumors to anticancer drugs and predict their efficacy have been developed [72]. Normal human LO models have been established to evaluate lung toxicity, and PHMG-p- or bleomycin-treated organoids showed fibrotic properties, such as increased expression of well-known epithelial-mesenchymal transition markers and extracellular matrix accumulation [59,73]. Overall, LOs have the advantage of recapitulating in vivo microarchitecture and function; thus, they are widely used as models of disease, development, drug screening, and toxicity evaluation.
SGs as a toxicity marker
In 2011, unidentified lung diseases involving severe respiratory distress were reported in a group of pregnant women who used humidifier disinfectants to prevent microbial contamination in Korea [74,75]. Numerous studies have reported that in vitro and in vivo exposure to PHMG, one of the components of humidifier disinfectants, induces inflammation and pulmonary fibrosis [76,77]. Due to the recent COVID-19 pandemic, the use of disinfectants, such as alcohol-based hand sanitizers and spray-type disinfectants, has increased significantly. Benzalkonium chloride (BZK), a widely used disinfectant with bactericidal properties, induces cytotoxicity in human lung epithelial cells depending on the exposure time and concentration [78].
In addition, BZK has been shown to induce lung inflammation and cellular damage, including interstitial edema, thickened alveolar septum, and decreased perivascular space in rats depending on the exposure concentration [79,80]. Biocides are often used in spray formulations to protect against microbial or viral contamination and to decontaminate surfaces, making respiratory exposure to biocides unavoidable [81]. Therefore, as concerns about the toxicity of hazardous substances such as hand sanitizers and biocides are increasing, the importance of accurate toxicity evaluation of biocides is being emphasized.
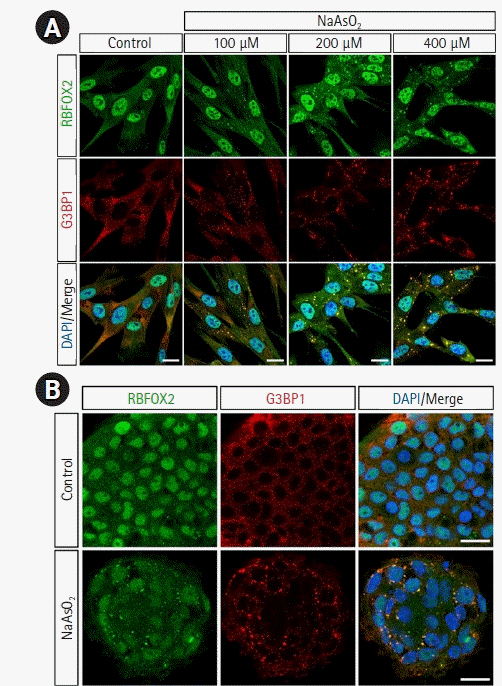
Several recent studies have suggested the possibility of exploring biomolecular condensates, such as SG formation, as important toxicity indicators. SG proteins, such as TIA-1 and PABP, are also localized in human cells when the cells are exposed to oxidative stress using NaAsO2 [82]. In 2017, Park et al. [83] made the novel suggestion that RBFOX2 is a component of SGs and plays an important role in regulating the mRNA and protein expression levels of retinoblastoma 1 when cells are exposed to stress. Furthermore, it has been reported that exposure to PHMG-p increases eIF2╬▒ phosphorylation in human LOs and BEAS-2B cells, and significantly induces SG formation under oxidative stress and viral infection conditions, resulting in the intracellular localization of G3BP1 and RBFOX2 [49]. In that study we showed the intracellular localization of RBFOX2 and G3BP1, the major components of SGs, in human pulmonary alveolar epithelial cells and human LOs after 1-hour treatment with NaAsO2 (Fig. 3). Therefore, based on the results of SG condensation in lung cells and LOs following exposure to (NaAsO2), we suggest that SG formation has the potential to be utilized as a toxicity marker for chemicals, including biocides.
Conclusion
Toxicity assessment of chemicals is becoming increasingly important in light of the occurrence of many deaths due to humidifier disinfectant exposure. In addition, concerns about respiratory diseases have increased significantly owing to the recent COVID-19 pandemic. In particular, as the use of disinfectants to prevent COVID-19 infection is rapidly increasing, there has been growing concern about whether the chemical substances in disinfectants are toxic to the respiratory system.
SG formation is a defense mechanism that protects cells from detrimental stress conditions, such as heat shock due to translational inhibition, oxidative stress, or viral infection. Unlike apoptosis, necrosis, and autophagy, SG formation is a dynamic and reversible process, in which SGs rapidly assemble under stress and disintegrate upon stress removal. However, long-term SG assembly can result in a persistent stress state, leading to cell death. Therefore, we suggest that SG formation can be used as a lung toxicity marker in the acute toxicity assessment of toxic and hazardous chemicals, such as disinfectants and biocides.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print