Introduction
The inner ear is a sensory organ with the physiological roles of balance and sound detection [1]. It is composed of two components with distinct functions: the cochlea, which detects sound and transmits it to the brain, and the vestibular system, which provides the sensation of balance. Damage to the inner ear, therefore, leads to impairment in sound detection and balance, causing hearing loss and vestibular disorders.
1. Hair cells
The inner ear is composed of various cells. Among them, hair cells play the most important role in the function of the inner ear. In the auditory system, sound energy passes through the auricle, the external ear canal, the eardrum, and the ossicles to create the movement of lymphatic fluid in the cochlea. In the vestibular system, lymphatic fluid movement in the utricle, the saccule, and the semicircular canals also occurs according to the body posture. Fluid flow is a type of mechanical movement that must be converted to chemical energy to transmit signals to the brain [2,3]. Hair cells are responsible for this conversion from mechanical to chemical energy in the inner ear.
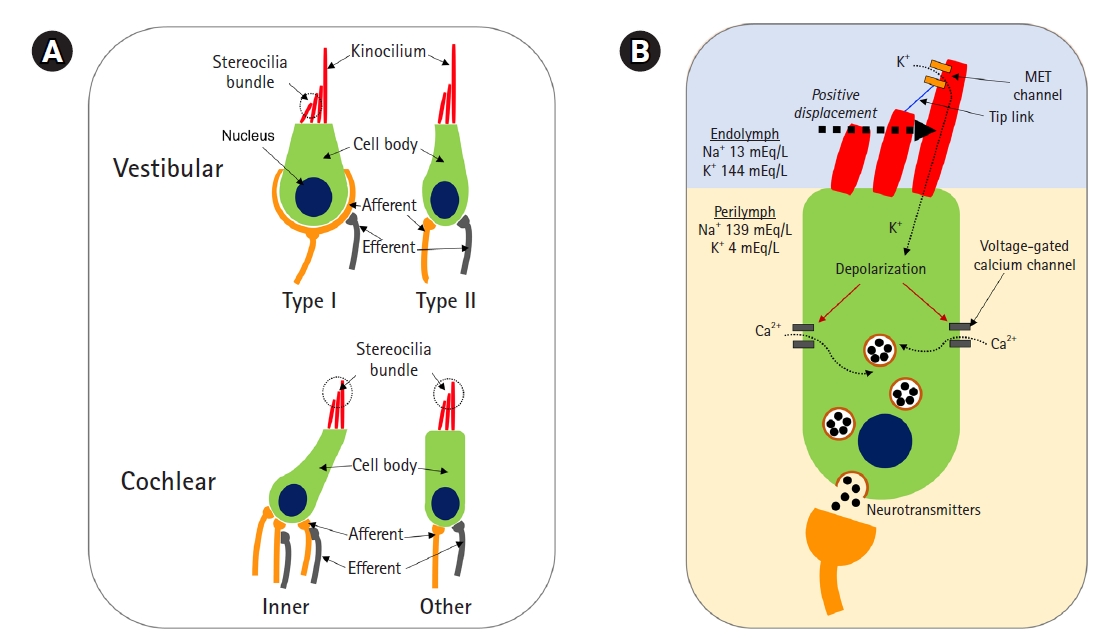
In the inner ear, there are four types of hair cells (Fig. 1A): inner hair cells (IHCs) and outer hair cells (OHCs) in the cochlea and type I and II hair cells in the vestibular system. Each has slight differences in its structure and function. The potassium current is the key to the function of those hair cells. For example, in the IHC, the mechanical movement of lymphatic fluid deflects the stereocilia, opening the mechanoelectrical transduction (MET) channel located in the stereocilia. The MET channel then permits potassium influx, and the IHC is depolarized (Fig. 1B) [4,5]. When the IHC is depolarized, the voltage-gated calcium channel is opened. Next, neurotransmitters are secreted, and auditory nerve fibers are activated [5,6].
Disorders related to inner ear dysfunction, including hearing loss, are mostly caused by damaged hair cells [7]. Of particular note, in mammals, the damage is permanent; hair cells neither regenerate nor recover except in very limited situations. Therefore, research on the inner ear focuses on those hair cells, including the prevention of cell damage and cell regeneration.
2. Research on hearing loss and vestibular disorders
Researchers in this field are more interested in cochlear hair cells (IHCs and OHCs) than in vestibular hair cells (type I and type II hair cells). This is because patients with disequilibrium can recover their balance functionality through vestibular compensation. In fact, balance involves integrating information from vision, proprioception, and the vestibular system. If one function (e.g., the vestibular system) is weakened, the others (e.g., vision and proprioception) are strengthened; as a result, the function of balance can be clinically restored by comprehensive vestibular rehabilitation therapy, allowing a patient to maintain a normal life [8-10].
In contrast, hearing loss resulting from damage to cochlear hair cells (IHCs and OHCs) does not recover except in very few cases. Thus, the prevalence of hearing loss is high; it is estimated to be 15% in all ages and reaches two-thirds in the older (≥70 years) population [11,12]. Furthermore, there is no medical treatment other than using devices, such as hearing aids and cochlear implants (CIs). Thus, in response to a high demand for treatment, research that ultimately aims at the development of drugs targeting hearing loss is actively underway worldwide [13]. Otic organoids are attracting attention because they can be used both as a platform for such research and as cell therapy to promote hair cell regeneration.
3. The organ of Corti
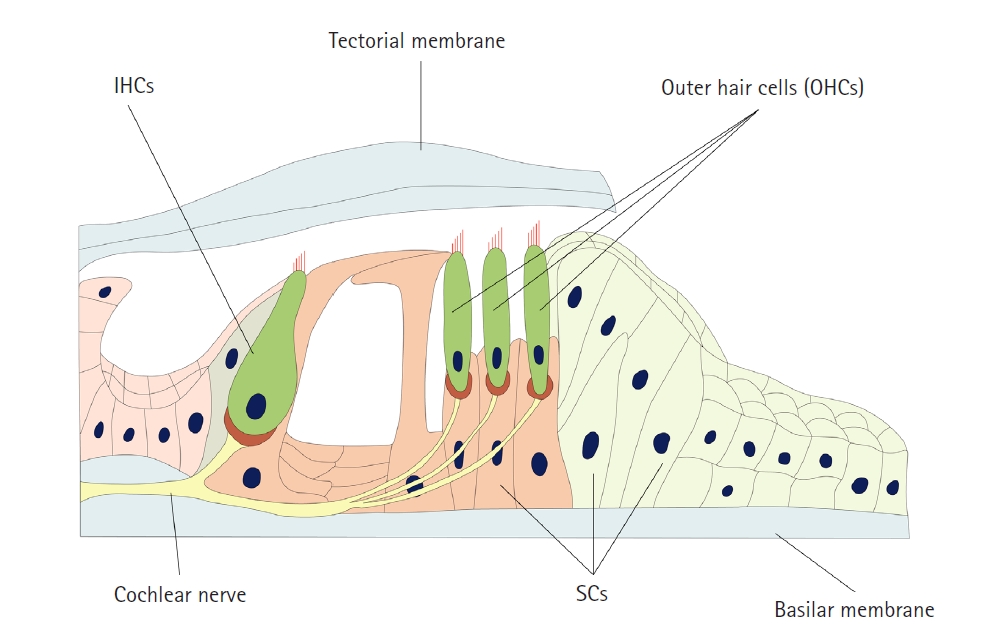
The cochlea is the organ responsible for auditory function. The cochlea is made up of IHCs, OHCs, supporting cells, the tectorial membrane, the basilar membrane, and the spiral ganglion, and those components maintain a structure called the organ of Corti (Fig. 2). This structure—the organ of Corti—plays a key role in hair cell function. The stereocilia of hair cells are contained in the endolymph, which maintains a high concentration of potassium. These cells can be depolarized by the influx of potassium through MET channels. Hair cells synapse with the spiral ganglion, and finally, auditory information is transmitted from the organ of Corti to the brain via the spiral ganglion [5].
4. Otic organoids
To summarize, the key elements in the production of otic organoids are as follows: (1) Differentiation of hair cell-like cells with structural similarities, including stereocilia; (2) Maturation of hair cell-like cells with functional similarities, including ion channels capable of potassium influx; (3) Overall, differentiation and maturation into cochlear hair cells, not vestibular hair cells; (4) Materialization of the organ of Corti, including hair cells, supporting cells, and the extracellular fluid with a high concentration of potassium.
Based on the above issues, this review article will discuss the development and progress of otic organoids, including future directions of research.
Development of otic organoids
1. Hair cell regeneration: transdifferentiation from supporting cells and differentiation from pluripotent stem cells
Researchers are interested in hair cell regeneration because most cases of hearing loss are caused by the permanent loss of hair cells. While hair cells cannot be regenerated in mammals, birds and amphibians can regenerate hair cells through two mechanisms: supporting cells may be transformed into hair cells, or they can divide to produce hair cell progenitors [14-16]. Therefore, extensive research has sought to determine how to transdifferentiate supporting cells into hair cells in mammals, and it is still actively underway [17-20].
Hair cell regeneration via transdifferentiation is expected to develop further in the future; however, it also has limitations. There are restrictions in terms of cell therapy that transplants those regenerated cells. For instance, gathering inner ear tissue from patients is not practically feasible, making it impossible to regenerate hair cells containing the genetic information of the patient via transdifferentiation.
Naturally, researchers have studied hair cell differentiation from pluripotent stem cells (PSCs). Initial experiments were based on a two-dimensional (2D) culture system used to generate retinal cells from PSCs [21]. Studies have demonstrated that hair cell-like cells with characteristics of hair cells could be differentiated from mouse and human PSCs [22-27]. However, since the above studies conducted 2D culture in Petri dishes and mainly focused on hair cells only, those cannot be considered as otic organoids.
2. Three-dimensional otic organoids
In 2013, Koehler et al. [28] reported that inner ear sensory epithelia could be differentiated from mouse PSCs using three-dimensional (3D) culture. The inner ear epithelia showed a specific structure similar to the inner ear, where hair cells and supporting cells maintain their own arrangement; these were referred to as the first otic organoids. Furthermore, in 2017, the same research team succeeded in differentiating otic organoids from human PSCs [29]. These researchers are the most advanced group in the development of otic organoids, and the protocols they presented are widely used in the generation of either mouse or human otic organoids [30,31].
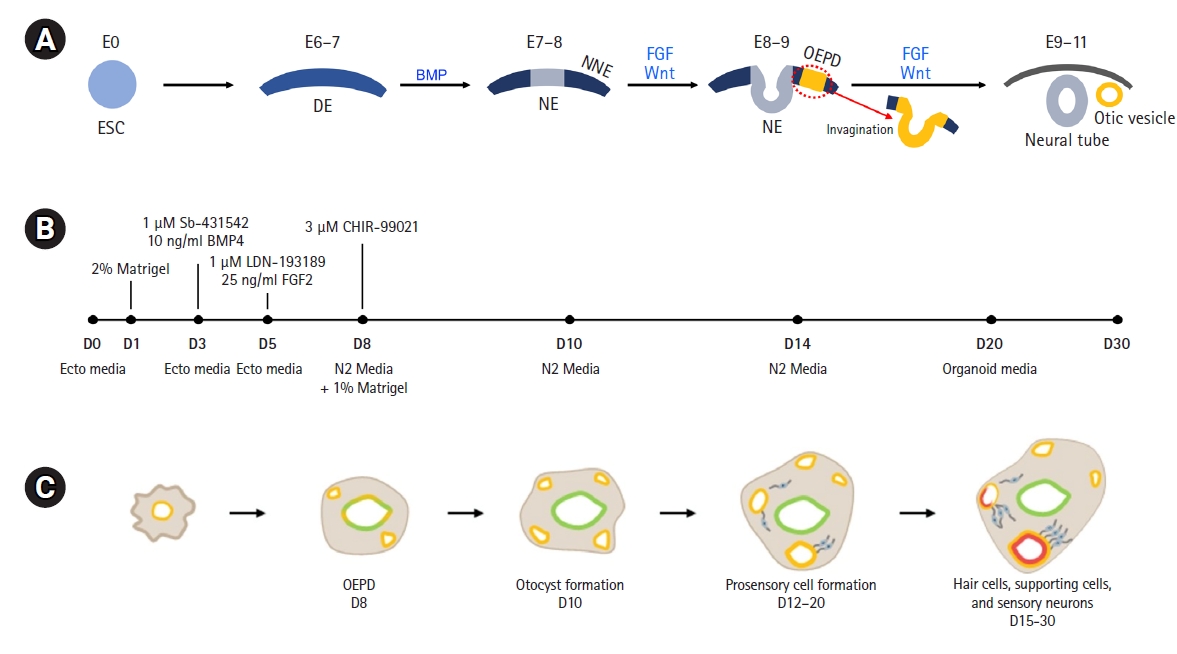
The aim of these protocols is to replicate the intricate and complex natural development process of the inner ear. Prior to a summary of the protocols, we provide a concise overview of the inner ear’s natural development process. The inner ear is derived from the ectoderm. It proceeds through the definite ectoderm (mouse embryonic day [E], 6-7), non-neural ectoderm (NNE) (E7-8), and preplacodal ectoderm [E8], and then the otic-epibranchial placode domain (OEPD) (E8-9) is formed, entering the otic fate. The OEPD invaginates from the surface of the ectoderm to create the otic pit, and then it breaks away from the ectodermal layer to form the otic vesicle (or otocyst). In this process of inner ear development, bone morphogenetic proteins (BMPs), fibroblast growth factors (FGFs), and Wnt signaling are known to play key roles (Fig. 3A) [32,33]. The protocol developed by Koehler et al. [28,30] follows the differentiation process of the inner ear, treating cells with these signal regulators to induce the formation of otic vesicles.
A brief summary of the generation of otic organoids is as follows. Using a 3D floating culture with 2% Matrigel as the extracellular matrix, the first step is to form PSC aggregates in a U-shaped-bottom microplate. Each U-shaped well has 3,000 to 5,000 PSCs. BMP4 for differentiation into NNE and a transforming growth factor-beta (TGF-β) inhibitor (SB-431542) for preventing mesoderm differentiation are administered on in vitro days 3 to 4. This step is critical in distinguishing the generation of otic organoids from other organoids related to the central nervous system. BMPs play a vital role in promoting differentiation into NNE, while inhibiting differentiation into neural ectoderm destined for the central nervous system [30]. On day 5, FGF2 and a BMP inhibitor (LDN-193189) are added to the medium; FGF2 plays a role in driving it towards the OEPD, whereas the BMP inhibitor prevents it from becoming epidermis. On day 8, a Wnt agonist (CHIR-99021) is administered to accelerate otic vesicle formation (Fig. 3B) [28-30]. An otic organoid containing hair cells, supporting cells, and sensory neurons forms in the vesicle. Mouse PSCs take 20 to 30 days for organoid formation, while up to 90 days are required for human PSCs (Fig. 3C).
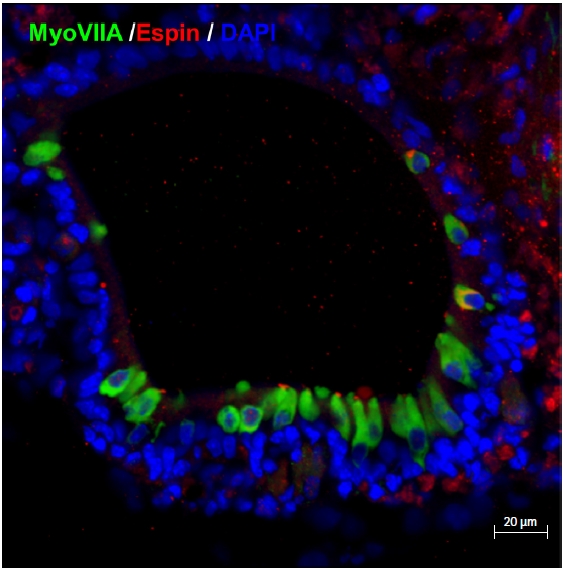
Starting on approximately day 10, these vesicles begin to form around the outer layer of a cell aggregate (Fig. 4). For the next 10 days, the sensory epithelia develop inside the vesicle. From day 20, the alignment of hair cell-like cells and supporting cell-like cells is identified in immunohistochemical staining (Fig. 5). Fig. 5 shows the most typical form of an otic organoid. Some of the immunohistochemical markers utilized for the characterization of cells in otic organoids include Sox2 (for the sensory epithelium including hair cells and supporting cells), MyoVIIa (for the cytosol of hair cells), Brn3c (for the nuclei of hair cells), α-tubulin (for kinocilia), Espin (for stereocilia), βIII-tubulin (for neurons), synaptophysin (for synapses), and Ctbp2 (for ribbon synapses) (for a comprehensive list, refer to Table 1 in the publication by Koehler and Hashino, 2014) [30]. Hair cell-like cells also have stereocilia. In addition, domains related to the synapse with nerves are observed. Although they are somewhat immature, it has been identified that the stereocilia of those hair cell-like cells are not only morphologically differentiated, but also function as ion channels.
Despite significant progress in the development of otic organoids, much remains to be elucidated regarding the incidental structures that may arise during this process. While nerve fiber-like structures have been frequently observed during otic organoid development, their significance beyond their synapse with the organoids remains unclear. Notably, a study has reported the spontaneous development of cardiac structures in some instances, which generate a bearing rhythm. It has been suggested that Wnt modulation may play a role in the formation of these unnecessary structures [34]. These findings underscore the need for continued refinement and improvement of otic organoid development protocols.
Applications of otic organoids in inner ear research
1. Current research platform in inner ear research
To date, hearing loss studies have used in vitro cell lines or in vivo animal models. For in vitro studies, a variety of immortalized cell lines have been developed, which have the characteristics of hair cells [35-37]. Among them, the HEI-OC1 cell line is well-known and is used as a screening tool before animal studies. Although novel research tools have been developed, the HEI-OC1 cell line continues to be widely used. However, we cannot say that HEI-OC1 cells exactly replicate any living beings. In addition, the hair cells are immature, with neither stereocilia nor MET channels [38].
The larval zebrafish lateral line is also used. It was first developed in 2005, and its hair cells are labeled with fluorescence [39]. Zebrafish are employed in studies elucidating the development of hair cells and the mechanisms of diseases, as well as for screening drug compounds [40,41]. The zebrafish lateral line is in vivo, making it superior to experiments using cell lines. However, all research should ultimately be applicable to humans, and the substantial inter-species differences between fish and mammals are the largest hurdle.
Therefore, animal studies still play a role in inner ear research. In addition to administering drugs directly to animals, we can also obtain cochlear explants from neonatal rodents; this method is referred to as ex vivo [42,43]. However, experiments using animals are increasingly facing skepticism; there are not only ethical issues, but also doubts about the effectiveness of the experiments [44,45]. Needless to say, a human experiment would be even more difficult to conduct.
2. Otic organoids as a research platform
Table 1 summarizes the characteristics of research methodologies. Experimentation using organoids may overcome the limitations of each method. Organoids would allow large-number studies because many organoids can be produced simultaneously. In addition, organoids generated from human PSCs naturally have a high similarity to humans. Another major advantage of this method is that it is free from ethical issues.
Due to the inability to biopsy the human inner ear without causing irreversible damage to hearing or balance, the study of human inner ear tissues has been restricted to scarce fetal and cadaveric tissue. Consequently, it has not been feasible to directly culture, infect, or treat inner ear tissue with toxic drugs, unlike other organs. However, the iPSC reprogramming and otic organoid generation technologies provide an exciting opportunity to study cultured tissue with the patient’s genetic information. This technology has enabled more extensive research on hereditary deafness and has already produced meaningful results [46]. A study on hearing loss caused by Tmprss3 mutations utilized otic organoids to elucidate the mechanism underlying the relationship between hearing loss and the mutation [47].
Otic organoids hold potential for investigating inner ear diseases that arise from infections. By directly infecting organoids with viruses or bacteria, the resultant effects can be studied. For example, a study infected otic organoids with COVID-19 to investigate its impact [48]. Additionally, organoids can serve as a platform for researching drug-induced ototoxicity, and the application of otic organoids has already commenced [49].
3. Otic organoids as cell therapy
Since there is no medical therapy for hearing loss, we cannot help but look forward to the therapeutic potential of otic organoids. In particular, there is some expectation that cell therapy might be possible; that is, that hair cells from organoids may replace the damaged cells of a patient. In fact, similar attempts have been made even before the generation of otic organoids. Mouse embryonic stem cells have been injected directly into the inner ear of an animal to determine whether those cells would differentiate into hair cells [50-55]. However, most cells could not be positioned appropriately. Although some cells could find the location—the endolymph space—they differentiated into neither hair cells nor neurons [50,52,53].
Some researchers attempted to transplant more differentiated cells, otic progenitor cells, into the inner ear [56-61]. The results were slightly better than when embryonic stem cells were transplanted; it was identified that those transplanted cells had synaptic connectivity with neurons. However, that finding did not provide sufficient support to attempt this transplantation for therapeutic purposes in humans. To sum up those attempts of cell therapy, at least, transplantation using a more differentiated cell state has a lower risk of teratoma formation and the cells are more likely to attach to the endolymph space [27]. Therefore, better results would be expected when highly differentiated cells from organoids are transplanted.
Current hurdles and future directions of otic organoids
As discussed above, otic organoids may lead to meaningful innovations in inner ear research, and significant progress has already been made. However, there remain many hurdles to overcome.
1. Differentiation into cochlear hair cells
Research on vestibular disorders is, of course, valuable and promising; however, in fact, researchers, clinicians, and patients have much higher expectations for the use of organoids in hearing loss studies. The cause of hearing loss is the loss of cochlear hair cells (IHCs or OHCs). Therefore, the production of otic organoids containing cochlear hair cells would be undoubtedly important for research on hearing loss. However, the properties of hair cells in otic organoids to date are closer to those of vestibular hair cells [28,29,62-64].
In a study using human PSCs, prestin, an OHC marker, was once identified, but except for that study [65], the types of hair cells were vestibular. Considering the inner ear developmental process, the default direction of differentiation after the otic vesicle is the vestibular system. The protocols to date do not contain any additional factors after vesicle formation, so this might be the natural outcome. This fact suggests that the otic organoids that have been generated to date are still immature. Although several efforts have been made for the maturation of otic organoids, a protocol for cochlear hair cell differentiation has not yet been established [34,67-69].
The protocol presented in Fig. 3 aims to promote differentiation up to the otocyst stage, but the actual process of inner ear development requires several important steps even after otocyst formation. Specific patterns of gene expression play a vital role in determining the subsequent stages of development, with two well-known types of patterning called the anterior-posterior (AP) axis and the dorsal-ventral (DV) axis [70]. Once these patterns are established, the development of specific inner ear components becomes possible. According to the AP axis, the neurosensory epithelium develops in the anterior domain of the otocyst while the non-sensory epithelium develops in the posterior domain, with retinoic acid and FGF8 playing essential roles in generating this pattern [71,72]. The DV patterning then determines whether the otocyst develops into the vestibular labyrinth (dorsal) or cochlea (ventral) [33]. Sonic hedgehog (SHH) is known to play a key role in this DV pattern, showing a gradient in which the concentration of SHH constantly increases from the dorsal to the ventral sides (Fig. 6) [73-75]. The current protocol for generating otic organoids does not utilize the signaling mediators mentioned above. To produce more developed otic organoids, it may be necessary to incorporate these additional regulators. However, the process may be complex, as it requires creating a gradient of the signal concentration.
Furthermore, in the process of its development, the inner ear must interact with the central nervous system. For example, SHH is secreted from the neural tube. However, the protocol we have used does not take into account this interaction with the surrounding structures. Therefore, the organoid proceeds toward the vestibular fate, which is the default, and it seems to be immature. As a result, it could be worthwhile to apply a co-culture system that can enable interactions between structures (Fig. 6) [76,77].
2. Yield efficiency and reproducibility in the generation of otic organoids
According to reports, otic organoids were formed in approximately 70% to 80% of all aggregates during differentiation with mouse PSCs, while the formation rate drops to around 20% when using human PSCs [29,64]. It is anticipated that this efficiency will improve over time, but accurately quantifying this efficiency remains challenging. Moreover, there is currently no agreement on how to evaluate the qualitative efficiency of otic organoids. Researchers are completely responsible for manually finding otic vesicles in an aggregate; without immunohistochemistry, it is not known whether a vesicle has been well differentiated. Identified organoids are not consistent in either the number or arrangement of cells.
In addition, the differentiation period is also a hurdle. Using mouse PSCs, an organoid could be completely formed within approximately 30 days, while the process takes 90 days with human PSCs. It is never easy to conduct organoid culture for 90 days without any unexpected problems in the laboratory. This efficiency issue should be solved for the industrialization of otic organoids. The current status might be disappointing, but this means that we should continue making progress until otic organoids can be used as a solid research methodology.
3. Inner ear organoid-on-a-chip
In the generation of otic organoids, it is worth considering the application of a microfluidic-based approach. The organ of Corti, a core structure of the cochlea, has not been developed. Using sophisticated 3D bioprinting to generate an inner ear organoid-on-a-chip, the organ of Corti may be simulated and the signaling gradient (e.g., the concentration gradient of SHH) can be implemented by controlling microfluids on the chip [78]. Investigations on microfluidic or microwell systems are being actively conducted, and these methods are expected to be utilized soon in otic organoids [79-81].
4. Otic organoids versus cochlear implantation
The auditory system in humans is the only sensory organ in our body, the function of which can be restored to the extent that our daily lives are possible via an implantable device—a CI. Cochlear hair cells convert mechanical energy from external and middle ear into electrochemical energy, and that is also what a CI does. Since CIs were first introduced in the 1980s, their effectiveness and safety have been fully demonstrated, and it has been confirmed that CIs can also relieve the social isolation caused by severe hearing loss; thus, the indications for CI surgery are gradually expanding [82-89]. According to the authors’ clinical experience of performing CI surgery and the studies we have conducted, there is no question about the effectiveness of CIs [90-95].
However, CIs cannot be a universal treatment applicable to all hearing loss patients because they require surgery and are indicated for severe hearing loss. The demand for drug treatment for hearing loss is still substantial. Furthermore, surgical experience with CIs will eventually be a major asset for organoid-based cell therapy. Because the electrode of a CI is inserted directly into the inner ear, the same approach can be used as a drug delivery modality (Fig. 7) [96-100]. This means that rapid progress might be possible in the field of otic organoid-based cell therapy.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print