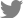Introduction
Fabry disease is a rare lipid storage disorder, but is more frequent within selected patient populations, such as those suffering from chronic kidney disease or hypertrophic cardiomyopathy [1-3]. Fabry disease results from an X-linked mutation in the α-galactosidase A (GLA) gene, producing GLA enzyme deficiency or absence. This leads to progressive lysosomal globotriaosylceramide (Gb3) accumulation and thus multi-organ damage and dysfunction (including renal, cerebrovascular, and cardiac) [4]. The vast majority of Fabry disease patients experience kidney damage, resulting in end-stage renal disease and early mortality [5]. While enzyme replacement therapy is available [6], it is extremely expensive and cannot reverse damage which has already occurred (e.g., in advanced disease) [7]. To receive enzyme replacement therapy treatment, Fabry disease patients must visit the hospital once every 2 weeks and receive intravenous injections for several hours. Oral treatments are being developed, and patient convenience is expected. However, there are hurdles to conducting effective evaluation of therapeutics. Because it is difficult to recruit sufficient numbers of patients for clinical trials, because no available in vivo animal model species fully recapitulates the human phenotype, and because no satisfactory predictive toxicology assay model currently exists [8]. Therefore, there is a critical need for an appropriate model that predictive high-throughput screening for drug development.
Organoids derived from pluripotent or adult stem cells are increasingly employed by various studies [9,10]. Given that organoids exhibit organ-like architecture and may recapitulate in vivo disease mechanisms, they are attractive tools for disease modelling [11]. While Itier et al. [12] developed a human induced pluripotent stem cell (hiPSC)-derived cardiomyocyte model of Fabry disease [13], no Fabry disease model based on nephron organoids differentiated from stem cells yet exists. The present study aimed to develop an organoid-based Fabry disease model derived from human embryonic stem cells (hESCs) featuring GLA mutations introduced via CRISPR/Cas9.
Materials and Methods
Ethics statement: The research described in this thesis utilized cell lines. As these cell lines were not directly obtained from human subjects, Institutional Review Board (IRB) approval was not sought. Research involving cell lines should consider the origin and ownership of the cell lines. The cell lines used in this study were provided by reputable suppliers, and their use was conducted in accordance with ethical guidelines and regulations.The authors of this thesis affirm that all research procedures involving the use of cell lines were conducted in compliance with ethical principles and guidelines, and efforts were made to ensure the accuracy and integrity of the research findings.
1. Cell culture
The CHA15 hESC line (CHA University, Korea) and H9 hESC line (WiCell, USA) were maintained on Matrigel (#354277; Corning) coated 35 mm dish (#150460; ThermoFisher) in mTeSR 1 (#85850; STEMCELL Technologies).
2. Cas9 protein purification and in vitro sgRNA transcription
Recombinant Cas9 protein was purchased from ToolGen Inc. (Seoul, Korea). Single guide RNAs (sgRNAs) were synthesized by in vitro transcription using T7 RNA polymerase (New England BioLabs). In brief, sgRNA templates were incubated with T7 RNA polymerase in reaction buffer (40 mM Tris-HCl, 20 mM MgCl2, 1 mM dithiothreitol (DTT), 2 mM spermidine, at pH 7.9; New England BioLabs), containing 4 mM NTPs (Jena Bioscience) and RNase inhibitor (New England BioLabs) for 16 hours at 37°C, followed by incubation with DNase I (New England BioLabs) for 30 minutes at 37°C. Synthesized sgRNAs were purified with a polymerase chain reaction (PCR) purification kit (Macrogen).
3. Cas9-ribonucleoprotein delivery
To edit GLA genes in hESCs, cells were transfected with a Cas9-ribonucleoprotein (RNP) complex using a 4D-Nucleofector (Lonza). The RNP complex was made by mixing 10 μg of Cas9 protein in storage buffer (20 mM HEPES pH 7.5, 150 mM KCl, 1 mM DTT, and 10% glycerol) with 7.5 μg of in vitro transcribed sgRNAs and then incubating the mixture for 15 minutes at room temperature (RT). We used 1×105 hESCs in 20 μL Primary P3 buffer and performed electroporation using the nucleofector program CB-150. After transfection, the cells were plated on Geltrex (Thermo Fisher Scientific)-coated 24-well plates and cultured in Essential 8TM Flex Medium containing RevitaCell Supplement (Thermo Fisher Scientific).
4. Nephron differentiation of hESC lines
hESCs grown on Matrigel were washed once with PBS (#LB001-02; WELGENE) and dissociated into single cells with Accutase (#07920; STEMCELL Technologies). Cells were then plated at a density of 1.6×104 (CHA-hES15) or 0.8×104 (H9) cells/cm2 onto 24-well tissue culture plates (#142475; ThermoFisher) coated with 2% Matrigel in mTeSR 1 supplemented with the ROCK inhibitor Y27632 (10 mM) (#1254; TOCRIS). After 72 hours, cells were briefly washed in phosphate-buffered saline (PBS) and then cultured in basic differentiation medium consisting of Advanced RPMI 1640 (#12633-020; ThermoFisher) and 1×GlutaMAX (#35050-061; ThermoFisher) supplemented with CHIR99021 (8 μM) (#4423; TOCRIS) for 4 days to induce late primitive streak cells. Noggin (5 ng/mL) (#CYT-600; PROSPEC) was also used for hiPSC differentiation in addition to CHIR (8 μM). To induce posterior intermediate mesoderm, cells were then cultured in Advanced RPMI+1×GlutaMAX+activin (10 ng/mL) (#338-AC-050; R&D Systems) for 3 days. For induction of nephron progenitor cells, the media was then changed to Advanced RPMI+1×GlutaMAX+FGF9 (10 ng/mL) (#273-F9-025/CF; R&D Systems) for 7 days. CHIR (3 mM) was added to the media from day 9 to 10 of differentiation to induce renal vesicles. On day 14, cells were switched to the basic differentiation medium and cultured for an additional 7 to 14 days (total of 21 to 28 days). The medium was replaced every 2 or 3 days.
5. Immunohistochemistry for kidney organoid analysis
The nephron organoids were washed once with PBS and fixed in 4% paraformaldehyde for 15 minutes at RT. Fixed organoids were washed 3 times in PBS and incubated in blocking buffer (0.3% Triton X-100 and 5% normal donkey serum) for 1 hour at RT. The organoids were then incubated with primary antibody overnight at 4°C in antibody dilution buffer (0.3% Triton X-100 and 1% bovine serum albumin in PBS). Cells were then washed 3 times in PBS and incubated with Alexa Fluor 488, 594 conjugated secondary antibodies (1:500) (Life Technologies) in antibody dilution buffer for 1 hour at RT. For primary antibody immunostaining with Lotus Tetragonolobus Lectin (LTL) (#FL-1321-2; Vector Labs), Podocalyxin (#AF1658; R&D Systems), E-Cadherin (#AF748; R&D Systems), N-cadherin (#ab12221; Abcam), alpha Galactosidase (#PA5-13687; ThermoFisher), Anti-Gb3 (#A2506; TCI). Nuclei were counterstained with Hoechst 33342 (#14533; Sigma).
6. Transmission electron microscopes
To observe the zebra body in Fabry’s disease nephron organoids, transmission electron microscope (TEM) analysis was performed. Briefly, Fabry’s disease nephron organoids were fixed in Karnovsky’s fixative (2% glutaraldehyde, 2% paraformaldehyde, 0.5% CaCl2 in 0.1 M phosphate buffer, pH 7.4), and embedded in the epoxy resin after post-fixation with 1% OsO4 dissolved in 0.1 M phosphate buffer. Ultrathin sections were processed using an ultramicrotome (EM-UC-7; Leica) and placed on a copper grid stained with 6% uranyl and lead citrate. The samples were observed using a JEM-1011 (JEOL).
Results
1. Successful GLA knockout in hESCs
Targeted indel mutations were introduced into GLA using the CRISPR/Cas9 genome-editing system (Fig. 1A). Nephron organoids were produced using GLA knock-out hESC. This was generated by the Fabry disease organoid model. Deep sequencing of genomic DNA from homozygous GLA-mutant hESCs confirmed the presence of intended mutations at the target site (Fig. 1B). Real-time quantitative PCR analysis confirmed that GLA was not expressed in the GLA knockout hESCs group compared to wild-type group (Fig. 1C).
2. Confirmation of hESC and Fabry disease phenotypes
Immunocytochemistry targeting hESC markers (POU domain, class 5 transcription factor 1 [Oct-4]; transcription factor SOX-2 (SOX-2); and homeobox protein NANOG [hNanog]), and Fabry disease phenotype-relevant GLA demonstrated: no difference in hESC marker expression between wild-type and GLA knockout hESCs (confirming preservation of hESC identity), and absent GLA expression levels only in GLA knockout hESCs (Fig. 1D).
3. Generation of kidney organoids and confirmation of nephron characteristics
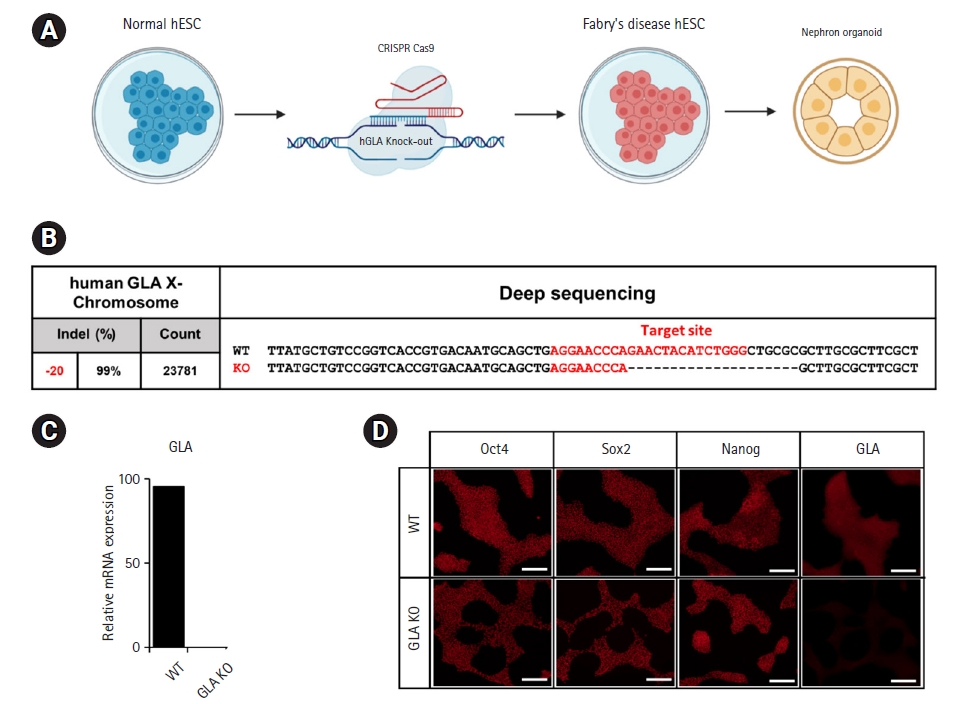
Both wild-type and GLA knockout hESCs were used to generate kidney organoids by means of sequential exposure to established differentiation conditions [14,15]. On differentiation day 21, renal vesicles formed nephron-like structures and expressed nephron-specific markers, including a podocyte marker (podocalyxin) and LTL-binding sites characteristic of proximal tubule epithelia (Fig. 2A and 2B). That these observations did not differ between wild-type and GLA knockout organoids demonstrates that GLA knockout did not alter the capacity of hESCs to differentiate into nephron-like structures.
4. Confirmation of organoid Fabry disease phenotype
When diagnosing Fabry disease, ultrastructural images (electron microscopy) of the glomeruli are examined to identify layered lipid inclusions (zebra bodies) within the podocyte cytoplasm. In this study, when GLA knockout organoids on day 21 of differentiation were observed by TEM, we observed the presence of zebra bodies within the organoids (Fig. 2C). In order to confirm the deficiency of GLA, which is a characteristic of Fabry disease, immunohistochemical staining was performed and it was confirmed that GLA was not expressed in the organoid disease model (Fig. 2D). Lysosomal accumulation of Gb3 is the element characterizing Fabry disease due to a hereditary deficiency GLA enzyme. Immunohistochemistry results showed that Gb3 accumulation of Fabry disease modelling organoid as contrasted with wild-type organoid (Fig. 2E). These findings indicate that GLA knockout hESC-derived kidney organoids imitative the key features observed in Fabry disease patients.
Discussion
Disease modelling employing human-derived organoid cultures is beneficial for both disease mechanism elucidation and drug screening [16]. Furthermore, this approach is extremely valuable in the investigation of rare diseases such as Fabry disease, as the limited number of available patients hampers drug evaluation.
Recent advances in the generation of kidney organoids, together with gene editing systems such as CRISPR/Cas9, have extended this approach to renal disease [17]. One example is an in vitro polycystic kidney disease model derived via differentiation of hPSCs featuring targeted PKD1 and PKD2 gene mutations. Similarly, Freedman et al. [18] used the CRISPR/Cas9 system to introduce appropriate targeted mutations, followed by differentiation of mutated cells into kidney organoids which recapitulate the renal tubule cell cysts characteristic of renal cystic disease. Among the organs affected during Fabry disease, the kidney is particularly significant: renal failure is the major cause of death. Although many studies have attempted to generate in vitro and in vivo models of Fabry disease, none have developed kidney organoid-based models. For example, Pereira et al. [19] generated a Fabry disease model based on CRISPR/Cas9-modified immortalized podocytes, and Itier et al. [12] differentiated patient-derived iPSCs into cardiomyocytes recapitulating lysosomal Gb3 accumulation.
The present study differentiated GLA knockout hESCs to successfully develop a kidney organoid-based Fabry disease model, in which organoids form nephron-like structures expressing podocyte- and proximal tubule epithelium-specific markers and recapitulate absent GLA expression concomitant with lysosomal Gb3 accumulation and Fabry disease-pathognomonic zebra body formation. Because the Fabry disease organoid model created through this study mimics the characteristics of human diseases, it can be used for screening new drug candidates or assessing the effectiveness of therapeutic agents. To the best of our knowledge, this is the first study to report that GLA knockout hESC-derived kidney organoids recapitulate Gb3 accumulation. To further validate model robustness for drug screening, it would be necessary to investigate whether currently available interventions (e.g., GLA replacement therapy) decrease Gb3 accumulation.
In conclusion, we report the first kidney organoid-based Fabry disease model, which may provide a cornerstone for further disease mechanism elucidation and therapeutic drug discovery.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print